Last Update Oct 31, 2010
研 究 内 容
(1) 森林の更新動態と種多様性維持機構の解明
(1-1) トドマツ・アカエゾマツ林の構造と2種の共存に関する研究
(1990〜1996,2006〜現在)
(1-2) 北海道の針広混交林の更新動態に関する研究 (1994〜一応,現在)
(1-3) ロシア・カムチャッカ半島の針広混交林の構造と更新に関する研究 (1998〜1999)
(2) 樹形の構造と機能に関する研究 (1991〜現在)
(3) ササが樹木の生長に及ぼす影響 (1998〜2000)
(4) 標高傾度にそった森林の更新動態(2000〜現在)
(5) カナダ東部の分布北限の落葉広葉樹林の更新動態(2005〜2010)
(6) 外来植物の生態系への影響 (2007〜現在)
(1) 森林の更新動態と種多様性維持機構の解明
複数の種がなぜ同じ場所で共存していられるのか? 一つの森林は通常,多くの種によって構成されており,それぞれの種が種子の発芽定着,その後の成長と死
亡という一連の生活史の過程を経て更新します.そして,これらの過程は種ごとに独立に行われることもあれば,種間競争などの生物間の相互作用も時には関係
することもあります.したがって,森林がどのように維持更新しているのか,そして種多様性がどのようにして保たれているのかを明らかにするために,生活史
の各過程から調べていく必要があります.今まで北海道の亜高山帯針葉樹林や針広混交林,そしてロシア・カムチャッカ半島の針広混交林について調べてきまし
た.
(1-1) トドマツ・アカエゾマツ林の構造と2種の共存に関する研究 (1990〜1996, 2006〜現在)
北海道の亜高山帯林ではしばしばトドマツとアカエゾマツの針葉樹2種が優占してます.この2種の共存機構について卒論から研究をはじめ,最後は博士論文と
してまとめました.2種の共存は2種間での競争だけで決まるわけではなく,発芽定着に利用する微地形と林床で優占するササとの関係が2種の共存に大きく影
響していることを明らかにしました.

雌阿寒岳の麓に成立したアカエゾマツ一斉林.
|

ササはなく,トドマツが侵入し,遷移が進行している. |
|

ササが適度にある温泉山(糠平).
ここでは2種がほどよく混交している
|

ササが非常に深い十勝岳.
アカエゾマツが優占している |
|
(1-2) 北海道の針広混交林の更新動態に関する研究 (1994〜一応,現在)
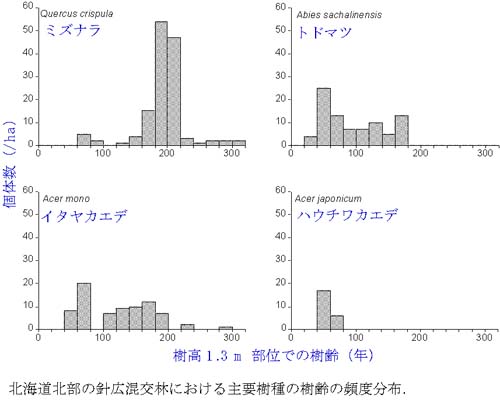
亜高山帯では針葉樹が優占していますが,比較的標高の低い場所では,針葉樹と広葉樹が混交した針広混交林が形成されます.北海道北部の針広混交林では,針
葉樹のトドマツとともに,ミズナラ,イタヤカエデ,ハウチワカエデといった広葉樹が分布しています.この森林の樹齢やサイズ構造を調べた結果,約200年
ほど前にミズナラが一斉に更新し,その後,トドマツ,イタヤカエデなど他の樹木が更新してきたことが分かりました.ミズナラの一斉更新の原因は解明できま
せんでしたが,針広混交林の更新には何らかの大きな攪乱が重要であることを示しています.北海道東部の針広混交林では針葉樹のトドマツが撹乱後に一斉に更
新している傾向が認められます.
(1-3) ロシア・カムチャッカ半島の針広混交林の構造と更新に関する研究 (1998〜1999)
ソビエト連邦の崩壊によって,軍事的に重要拠点であったカムチャッカ半島も外国人に開放されるようになりました.カムチャッカ半島の中央は両側を山脈には
さまれた低地帯になっています.そのため降水量が少なく,植生も他の地域と異なっています.カムチャッカ半島全体ではダケカンバがもっとも優占しています
が,中央低地帯ではシラカンバやエゾマツが優占しています.この森林の構造を調べた結果,シラカンバはおもに萌芽によって更新し,そしてエゾマツはおもに
親木の下で発芽定着していました.その結果,両種は排他的に分布しているため,種間競争がほとんど生じることなく,共存していることが分かりました.エゾ
マツの実生の定着が親木の下に制限されるのは,乾燥気候のため,暗い親木の下は乾燥害が低いためと思われます.このようにカムチャッカ半島の森林の更新は
気象条件に大きく影響されていることが示唆されました.

エゾマツ・シラカバが優占した針広混交林.
|

カレイタ氷河の後退跡地で植生遷移を調べました. |
|
(2) 樹形の構造と機能に関する研究 (1991〜現在)
樹形とは文字どおり木の形のことです.特に枝と葉で構成されている樹冠の部分を指します.一本の木には多くの枝があり,そしてそれらがまとまって樹冠を形
成します.樹形は一見,なんの秩序もなく形成されているように見えます.しかし,樹形とは樹木の更新,あるいは成長や生存戦略に大きく影響する重要なもの
なのです.特に下層個体は暗い林冠下で生存しなければならず,また環境が成長に適した明るい環境になれば,すばやく成長する必要があります.私は,このよ
うな下層個体の樹形の構築様式がどのようにその種の成長戦略に反映されているのかについて興味があります.今まで,針葉樹や落葉広葉樹,そして熱帯雨林の
ショウガ科植物,林床性ヤシなど,幅広い生活形について研究してきました.

サトウカエデの実生.葉が重ならないように,
葉柄で葉の配置がうまく調節されている.
|

インドネシアの熱帯雨林での調査風景.
樹形の測定と葉のマーキングをしているところ. |
|
(3) ササが樹木の生長に及ぼす影響 (1998〜2000)
日本の森林の特徴は林床がササで被われていることです.ササの存在によって林床は暗くなり,そのため樹木の実生の生存・成長率は低下し,結果的にササに
よって森林の更新は妨げられます.今までのササに関する研究は,このようなササの実生(つまりササよりも小さな個体)への影響に関する研究がほとんどでし
た.ところが,ササの樹木への影響はそれだけではないはずです.地下部の競争をとおして,ササよりも大きな個体の成長に影響している可能性もあるはずで
す.大規模なササの除去実験によって,この可能性を示すことができました.
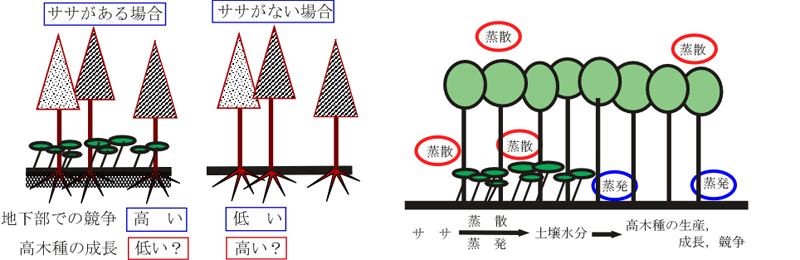
高さ2mのチシマザサを刈り払った調査区.自分で言うのもなんですが,かなり,たいへんな作業でした.
(4) 標高傾度にそった森林の更新動態(2000〜現在)
現在,化石燃料の使用の増加によって地球温暖化が進行している.地球温暖化は植生分布にどのような影響をもたらすのだろうか? 標高が上がるにしたがい気
温は低下し,植生が変化する.例えば,中部山岳地域では植生は落葉広葉樹林から針葉樹林,そしてハイマツ林へと変化する.このような植生の変化は温度条件
と関係していることは疑いがない.では,地球温暖化は標高傾度にそった現在の植生分布をさらに上の標高まで押し上げるのだろうか? この問いに答えること
は難しいでしょう.まず,標高傾度にそった現在の植生分布が気象条件とどのように関係しているのかを明かにする必要がある.そのために,標高傾度にそって
「植生と森林構造」,「成長と気象の関係」や「森林限界の形成機構」などを調査している.とくに,成長と気象の関係については同一の標高帯でも種によって
気象条件に対する反応は若干異なるが,低い標高ほど乾燥ストレス,高い標高ほど日照不足や低温ストレスが大きくなっている傾向が見いだされている.

森林限界では風雪のため,樹木は正常には
成長できない.
|
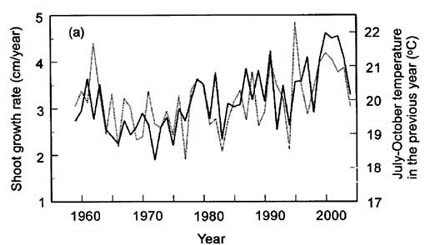
ハイマツ(実線)の成長は前年夏の気温(点線)と強い相関がある. |
|
(5) カナダ東部の分布北限の落葉広葉樹林の更新動態(2005〜2010)
モントリオールのあたりは落葉広葉樹林の分布北限となっている.このあたりでは,夏季は温暖な気候であるが,冬季はしばしば ice storm
が発生し,森林に大きな被害を及ぼす.また,分布北限での更新動態は他の分布域とは異なると考えられる.そこで,モントリオールから車で一時間ほどのとこ
ろにある Mont St. Hilaire において,サトウカエデとアメリカブナの優占した old-growth forest
の更新動態を気象条件との関係,そして稚樹の光環境に対する応答などを調べている.

MSHのピークからの風景.
|

Old-growth beech-maple forest. |
|
(6) 外来植物の生態系への影響 (2007〜現在)
ヒトやモノの移動にともない,多くの生物種が本来の生息地から離れた場所に分布するようになった.代表的なものは国境を越えてくる外来植物であるが,国内
でも,ある生態系から別の生態系の侵入,移動もこの問題に含まれる.それぞれの生態系は多くの種から構成されているが,それは長い時間をかけて種間関係が
成立してきた.そのため,外来種の移入はその場所の固有の生態系に大きなインパクトを与えると考えられる.そこで,手始めとして,日本の河川敷に広く分布
しているビロー
ドモウズイカ(地中海地方原産)の現状把握とその個体群維持機構の調査を2007年から始めた.

戻 る

